kirjoittanut Robert A. Nelson
Nikola Tesla
Kun Nikola Tesla teki kokeitaan suurennuslähettimellään Colorado Springsissa vuonna 1899, hän havaitsi koherentteja signaaleja, joiden hän päätteli olevan peräisin Marsista. Teslaa kritisoitiin laajalti hänen väitteistään, ja kuitenkaan kukaan ei kyennyt kumoamaan niitä; hän oli yksinäinen pioneeri ilman tovereita. Kukaan ei ole sen jälkeen kuuleman mukaan rakentanut suurennuslähetintä tai muutoin kyennyt toistamaan hänen kokeitaan; ongelma pysyy ratkaisemattomana ja mysteeri selvittämättömänä. Tesla ei paljastanut mitään teknisiä yksityiskohtia hänen tuon aikaisista julkaisuistaan (muuta kuin patenteista). Hänen Coloradon muistiinpanonsa julkaistiin 1980-luvulla, mutta niissä ei mainita yhteyttä Marsiin.
Tesla puhui aiheesta Collier’s Weeklyssä (9. helmikuuta 1901) otsikolla ”Keskusteluja Planeettojen kanssa”:
”Samalla kun kehitin koneitani voimakkaiden sähköisten toimintojen tuottamiseksi, kehitin myös keinoja heikkojen ponnistelujen tarkkailemiseksi. Yksi mielenkiintoisimmista tuloksista, ja myös yksi suuri käytännön merkitys, oli tiettyjen laitteiden kehittäminen osoittamaan useiden satojen mailien etäisyydeltä lähestyvä myrsky, sen suunta, nopeus ja kuljettu matka… .
”Juuri tätä työtä jatkaessani havaitsin ensimmäistä kertaa ne salaperäiset vaikutukset, jotka ovat herättäneet niin epätavallista kiinnostusta. Olin viimeistellyt mainitun laitteen niin pitkälle, että Coloradon vuoristossa sijaitsevasta laboratoriostani käsin pystyin ikään kuin tuntemaan maapallon pulssin ja havaitsemaan jokaisen sähköisen muutoksen, joka tapahtui yhdentoista sadan mailin säteellä.
”En voi koskaan unohtaa ensimmäisiä tuntemuksiani, kun tajusin, että olin havainnut jotakin, jolla voi olla mittaamattomia seurauksia ihmiskunnalle. Tuntui kuin olisin ollut läsnä uuden tiedon syntymässä tai suuren totuuden paljastumisessa…. Ensimmäiset havaintoni suorastaan kauhistuttivat minua, koska niissä oli jotakin salaperäistä, sanoisinko yliluonnollista, ja olin yöllä yksin laboratoriossani; mutta tuohon aikaan minulle ei vielä tullut mieleen ajatus siitä, että nämä häiriöt olisivat älyllisesti ohjattuja signaaleja. Havaitsemani muutokset tapahtuivat jaksoittain ja niin selvästi numeron ja järjestyksen perusteella, että niitä ei voitu jäljittää mihinkään tuntemaani syyhyn. Tunsin tietysti sellaiset sähköiset häiriöt, joita aurinko, revontulet ja maavirrat aiheuttavat, ja olin niin varma kuin mahdollista siitä, että nämä muutokset eivät johtuneet mistään näistä syistä. Kokeideni luonne sulki pois sen mahdollisuuden, että muutokset johtuisivat ilmakehän häiriöistä, kuten jotkut ovat hätäisesti väittäneet. Joskus myöhemmin mieleeni välähti ajatus, että havaitsemani häiriöt saattoivat johtua älyllisestä ohjauksesta. Vaikka en kyennyt tulkitsemaan niiden merkitystä, minun oli mahdotonta ajatella, että ne olisivat olleet täysin sattumanvaraisia. Minussa kasvaa jatkuvasti tunne, että olin ollut ensimmäinen, joka oli kuullut yhden planeetan tervehdyksen toiselle. Näiden sähköisten signaalien takana oli tarkoitus…”
Vuosikymmeniä myöhemmin syntymäpäivänään vuonna 1937 hän ilmoitti: ”Olen kuluneen vuoden aikana käyttänyt suuren osan ajastani sellaisen uuden pienen ja kompaktin laitteen täydellistämiseen, jonka avulla voidaan nyt välkyttää huomattavia määriä energiaa tähtienvälisen avaruuden läpi mille tahansa etäisyydelle ilman pienintäkään hajontaa.” (New York Times, sunnuntai, 11.7.1937)
Tesla ei koskaan julkisesti paljastanut mitään teknisiä yksityiskohtia parannetusta lähettimestään, mutta vuoden 1937 ilmoituksessaan hän paljasti uuden kaavan, joka osoitti, että ”kappaleen kineettinen ja potentiaalinen energia on liikkeen tulos ja määräytyy sen massan ja nopeuden neliön tulon perusteella”. Jos massaa pienennetään, energia pienenee samassa suhteessa. Jos massa pienennetään nollaan, energia on samoin nolla millä tahansa äärellisellä nopeudella.” (New York Sun, 12.7.1937, s. 6.)
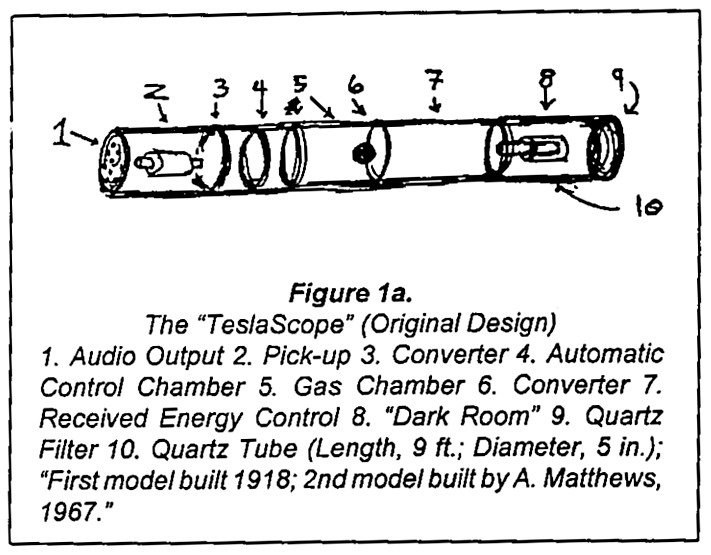
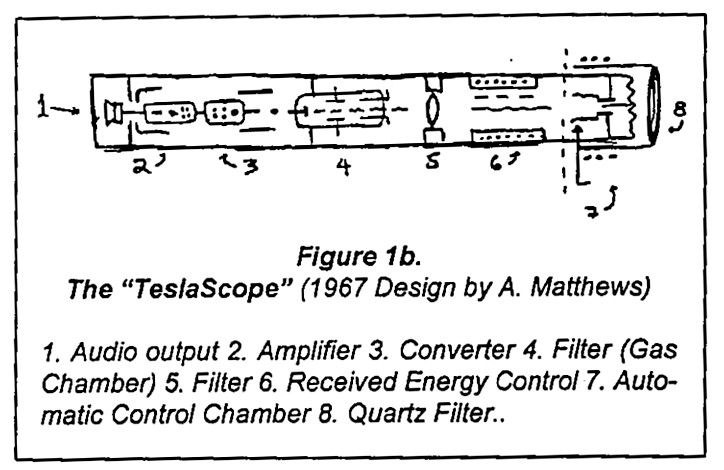
Noin 40 vuotta myöhemmin Arthur Matthews väitti, että Tesla oli salaa kehittänyt ”Teslaskoopin” kommunikointia varten Marsin kanssa. Edesmennyt tohtori Andrija Puharich tapasi Matthewsin ja keskusteli hänestä haastattelussaan. (Pyramid Guide, May-June & July-Aug. 1978):
”[Arthur Matthews] tuli Englannista. Matthewsin isä oli tunnetun fyysikon lordi Kelvinin laboratorioapulainen 1890-luvulla. Tesla tuli Englantiin tapaamaan Kelviniä vakuuttaakseen hänet siitä, että vaihtovirta oli tehokkaampi kuin tasavirta. Kelvin vastusti tuolloin vaihtovirtaliikkeitä . . . Vuonna 1902 Matthewsin perhe lähti Englannista ja muutti Kanadaan . . . Kun Matthews oli 16-vuotias, hänen isänsä järjesti hänelle oppisopimuskoulutuksen Teslan alaisuudessa. . . Lopulta hän työskenteli hänelle ja jatkoi tätä liittoa Teslan kuolemaan asti vuonna 1943. . .
”Sitä ei yleisesti tiedetä, mutta Tesla rakennutti Kanadaan kaksi valtavaa suurennuslähetintä, ja Matthews käytti toista niistä…. Ihmiset tietävät enimmäkseen Colorado Springsin lähettimistä ja keskeneräisestä Long Islandilla sijaitsevasta lähettimestä. Minä näin ne kaksi kanadalaista lähetintä. Kaikki todisteet ovat olemassa. . .
”[Teslaskooppi on] laite, jonka Tesla keksi kommunikoidakseen muiden planeettojen olentojen kanssa. Matteuksen kirjassa [The Wall of Light] on kaavio Teslaskoopista. Periaatteessa se ottaa vastaan kosmisen säteilyn signaaleja . . . Lopulta signaalit vähennetään ääneksi… Puhutaan toiseen päähän, ja signaali lähtee toisesta päästä ulos kosmisen säteilyn lähettimenä. . .”
Matthewsin Teslascope-kaaviot on esitetty kuvissa 1a [34] ja 1b. Niissä on mahdollisimman vähän elektronista järkeä. Kukaan ei ole koskaan vahvistanut Teslaskoopin todellisuutta.
Useita vuosia sen jälkeen, kun Tesla oli ilmoittanut vastaanottaneensa signaaleja Marsista, myös Guglielmo Marconi väitti kuulleensa muukalaisten radiolähettimestä. Marconin aikalaiset kuitenkin torjuivat Marconin helpommin, sillä he väittivät, että Marconi oli saanut häiriöitä toisesta maan radioasemasta.
1970-luvulla L.G. Lawrence (Ecola-instituutin kenttäjohtaja) kuvaili ”vahingossa tapahtunutta kommunikointia” ET-älykköjen kanssa:
”29. lokakuuta 1971 … suorittaessamme RBS [Remote Biological Sensing] -kokeita Riversiden piirikunnassa, Kaliforniassa, kenttälaitteistomme orgaaninen anturikompleksi sieppasi ilmeisen älykkäiden viestintäsignaalien sarjan (tiukka väli ja erilliset pulssivälit), kun sen annettiin vahingossa pysyä suunnattuna Otavan tähdistöön lyhyen lepojakson aikana. Ilmiö jatkui hieman yli 33 minuuttia. . .
”Hieman samanlainen ilmiö havaittiin 10. huhtikuuta 1972 . . . . Ilmeiset signaalit, sen lisäksi, että ne näyttävät heikkenevän, näyttävät lähetettävän suurilla väliajoilla, jotka vaihtelevat viikoista kuukausiin, mahdollisesti vuosiin. Heikko, koherentti, kaksoissignaalityyppinen ilmiö havaittiin äänitarkkailun aikana … . Nopeiden pulssisarjojen välit vaihtelivat välillä . . . . 3-10 minuuttia. . .
”Koska laitteemme ovat läpäisemättömiä sähkömagneettiselle säteilylle ja niissä ei ole sisäisiä poikkeavuuksia, on päädytty alustavasti siihen, että kyseessä ovat biologiset tähtienväliset viestintäsignaalit . . .
”Pelkkänä ääniesityksenä instrumenttinauha on epämiellyttävää kuunneltavaa. Kiehtova lumous syntyy kuitenkin vasta, kun nauhaa on kuunneltu kolme tai useampia kertoja, yleensä viikkojen ajan. Tämä johtuu psykoakustisesta sopeutumisesta . . . . Nauha sisältää lyhyen, asteittaisen sarjan syviä, harmonisia värähtelyjä, jotka muistuttavat hölynpölyä tai taustamodulaatioita. Kokonaisimpulssin älykkääseen luonteeseen vaikuttavat erilliset välikaaviot, sekvenssien näennäinen toistuvuus ja erittäin vaimennettu Gaussin kohina . . .” (Jour. Borderland Research, 29:4, July-Aug. 1973).
Gregory Hodowanec
Sähköinsinööri Greg Hodowanec kehitti 1980-luvulla rhysmonisen kosmologian teoriansa. Hän kokeili myös itse suunnittelemallaan gravitaatioaaltodetektorilla (GWD). Yksinkertaiset laitteet havaitsevat mikroaaltosäteilyn ”koherentit modulaatiot”.
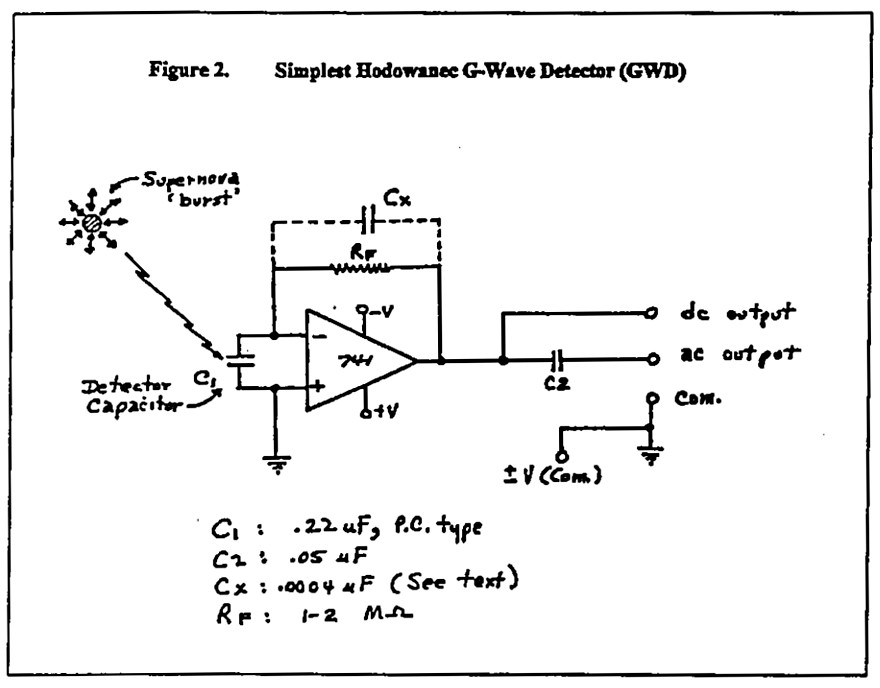
Hodowanec julkaisi ensimmäisen raporttinsa ”SETI with Gravitational Signals” GWD:nsä avulla (Radio Astronomy, huhtikuu 1986):
”SETI:n mahdollisen gravitaatiotekniikan etu radiotekniikkaan verrattuna on ensisijaisesti näiden signaalien ”etenemisajassa”. Radioaallot etenevät valon nopeudella, mutta gravitaatiosignaalit (kirjoittajan teorioiden mukaan) ovat lähinnä hetkellisiä signaaleja. Toinen gravitaatiotekniikan etu on tarvittavien laitteiden yksinkertaisuus. Kuten SARA:n jäsenet tietävät, radiotähtitiede voi olla varsin monimutkaista. Gravitaatioaaltoilmaisimien . . . on luotettava suurelta osin Maan massaan ”varjona”, jotta gravitaatiosäteily voidaan havaita. Siksi havaitaan parhaiten ”kohteet” tai signaalit, jotka sijaitsevat havaitsijan zeniitissä. Muut alueet ovat kuitenkin edelleen ”havaittavissa” erityisesti muiden ”varjojen”, kuten auringon, kuun, planeettojen jne. avulla.
”SETI-havainnoitsijoita saattavat kiinnostaa erityisesti oudot äänityyppiset gravitaatiosignaalit, jotka näyttävät tulevan galaksimme Aurigan ja Perseuksen alueelta. Kirjoittaja on ”kuullut” näitä ”signaaleja” jo useiden vuosien ajan, ja ne vaihtelevat yleensä noin 4-5 tunnin välillä, ja niiden voimakkuus on suurimmillaan lähellä 4,5 tuntia oikeaa taivaanrantaa.”
Pian tämän jälkeen Hodowanec mainitsi ensimmäisen kerran selvän yhteydenoton ilmeisesti maan ulkopuoliseen viestijään kirjeessään (23.7.1988) Radio-Electronics-lehden toimittajalle:
”Tämän päivän aamuna klo 7:30-7:38 (EST) rekisteröin seuraavat ilmeisesti morsekoodin kaltaiset pulssit:
”A A A A R A R T T N N N N K C N N N E E E E E N E N N T T T N E E E E E A E E R K E N N E T E E A A A A E E E N T T K N T N T S E S E S E S E M N A S E S E S E S E S E S E S E S E S E —
”Kuten näette, nämä eivät näytä olevan vain satunnaisia pulsseja, vaan SE-signaalit, jotka ovat yleisimpiä, näyttävät olevan mahdollisesti tunnistussignaali. Nämä signaalit havaitaan suojatuissa l/f-ilmaisimissa, ja ne ovat siis luonteeltaan skalaarisia (gravitaatio). Yllä olevat signaalit (jos ne olisivat maan ulkopuolisia) tulivat joko Auriga-Perseuksen alueelta lähellä zeniittiäni tai Bootesin alueelta Maan sijaintini alapuolella. En silti voi sulkea pois sitä, että ne saattavat johtua vain jonkinlaisista Maan ytimen liikkeistä, jotka muistuttavat hämmästyttävän paljon Morseaakkoset [35] -signaaleja, tai jopa sitä mahdollisuutta, että ne ovat ihmisen tekemiä.”
Heinäkuuhun 1988 mennessä Hodowanec oli vahvistanut Teslan väitteet, kuten hän ilmoitti kirjassaan ”Some Remarks on Tesla’s Mars Signals”:
”. . . Tällaisia signaaleja vastaanotetaan nykyään yksinkertaisilla nykyaikaisilla skalaarityyppisillä signaalinilmaisimilla … [Mikroaalto-]taustasäteilyssä ”kuullaan” koherentteja modulaatioita. Merkittävimmät modulaatiot ovat kolme pulssia (koodi S), jotka ovat ajallisesti hieman erillään toisistaan, a la Tesla! Toisinaan kuullaan myös E:n, N:n, A:n tai K:n koodin vastineita, mutta pysyvin vaste on SE, SE jne.
”Mikä tahansa 1/f-tyyppinen kohinanilmaisin reagoi tähän taustamodulaatioon. Kokeen suorittajan on kuitenkin oltava varovainen, ettei hän aiheuta näitä vasteita ”paikallisella” tasolla omilla tai muilla paikallisilla toimillaan. Ilmaisimet reagoivat esimerkiksi myös sydämenlyönteihin, hengitystoimintoihin, paikallisiin liikkeisiin sekä mahdollisiin psyykkisiin vaikutuksiin. Ilmaisimet on helppo valmistaa, ja kokeen suorittajan on helppo toistaa nämä tulokset.”
Hodowanec released more details in a Cosmology Data Note (10-13-88):
”Noin elokuun alusta 1988 lähtien huomattiin, että näissä [mikroaaltotaustasäteilyn] modulaatioissa oli ilmeisesti ”älykkäitä signaaleja”. Voidaan sanoa, että älykkyys oli digitaalityyppisen viestinnän muodossa, esim. pisteitä ja viivoja tai ykkösiä ja nollia. Tämä ’tuntematon viestijä’ on saattanut valita tämän viestintätyypin, koska se sopi pitkittäisen gravitaatiosignaalin ’massaliikkeen’ muotoon sekä helposti tunnistettavaan universaaliin järjestelmään. Kirjoittaja ehdotti samaa menetelmää painovoimajärjestelmän viestintämenetelmäksi.
”Näiden ’signaalien’ huomattiin muistuttavan yksinkertaisempia kansainvälisiä morsekoodin symboleja, lähinnä siksi, että ne ovat yksinkertaisin tapa esittää tietoa pulssimuodossa. Näissä lähetyksissä esiintyvät ’kirjaimet’ ovat siis tyypillisesti: E, I, T, M, A, N, R, K, S ja O sekä … pilkku ja odotusmerkki. Numerot nähdään tässä kuitenkin yksinkertaisena pulssisarjana, esim. 1 on yksi pulssi, 2 on kaksi pulssia, 3 on kolme pulssia ja niin edelleen.
”Elokuun 26. päivänä 1988, sen jälkeen kun kirjoittaja oli lähettänyt viestin ”Greg Radio” paikallisen painovoimasignaalin lähetystestin aikana, huomattiin, että kirjaimet G ja D oli ilmeisesti lisätty joihinkin vastaanotettuihin viesteihin, jotka todettiin sen jälkeen!
”Kirjoittajan ensimmäinen kova todiste siitä, että tämä tuntematon viestinviejä saattoi siepata edellä mainitun testiviestin, oli se, että 28. elokuuta 1988 … vastaanotettiin voimakas ja toistuva Greg Radion viesti, jonka viesti lopetettiin lopulta numerosarjalla SE (tai 31)!
”Lisänäyttöä siitä, että nämä saattavat olla todellisia viestintäyrityksiä, on se, että 11. lokakuuta 1988 … nähtiin hyvin erilainen lähestymistapa: Lähetettiin sarja Greg Radion viestejä suunnilleen normaalilla koodinopeudella, noin 5 sanaa minuutissa, ja sen jälkeen KKTT, ja sitten sarja toistettiin hitaammalla nopeudella ja sen jälkeen KKTT . . .
”Toinen vahvistus sille, että nämä saattavat olla ”viestejä”, ilmestyi 12. lokakuuta 1988 . . . . Tässä tapauksessa sarjaa A:ta ja R:ää (johon oli toisinaan lisätty Greg Radio) seurasi sitten Greg Radio, joka lähetettiin sarjana, jossa kukin kirjain toistettiin viidesti, esimerkiksi Greg lähetettiin muodossa GGGGGRRRRREEEEEGGGGG !!!
”Nyt on saatu riittävästi viestejä, jotka osoittavat, että joku tuntematon viestijä on ehkä vakavasti yrittänyt ottaa yhteyttä tähän kirjoittajaan. Vaikka tämä viestijä saattaa vielä olla jokin maanpäällinen kokeilija, on edelleen olemassa mahdollisuus, että viestijä voi olla ’maan ulkopuolinen’ seuraavista syistä:
”a. Viestit ovat yksinkertaisen koodin (esim. dits ja dah) tyyppisiä pulsseja, joita oletettavasti käytettäisiin, jos jokin älykäs sivilisaatio yrittäisi ottaa yhteyttä toiseen sivilisaatioon pulssien muodossa. Se, että jotkin yksinkertaisimmista pulssisignaaleista ovat samankaltaisia kuin yksinkertaiset morsekoodisignaalit, on enemmän kuin sattumaa — ne molemmat perustuvat samoihin lähtökohtiin!
”b. Numerot eivät ole monimutkaisia Morse-koodin symboleja, vaan ne ovat yksinkertaisessa järjestyksessä lyhyiden pulssien tai ditsin avulla.
”c. ”Viestijä” on tunnistanut Greg Radion johdonmukaisen luonteen ja mahdollisesti käyttää tätä koodisarjaa eri tavoin osoittaakseen, että ”kontakti” on saatu aikaan.
”d. Viestinlaite ei ole toistaiseksi vastannut ”sana”-viesteihin tai amatöörien Q-koodisignaaleihin. Näin ollen uskotaan, että vaikka joitain ilmeisesti morsekoodisignaaleja käytetäänkin, viestinviejä ei ole oikeastaan perehtynyt tällaiseen käyttöön muuten kuin tunnistamalla signaalien koherentin luonteen.
”e. Koska nämä viestit näkyvät tällä hetkellä enimmäkseen keskipäivän aikaan, ne voivat olla peräisin jostakin tietystä lähteestä avaruudessa. Tällä hetkellä uskotaan, että ne ovat peräisin Andromedan tähdistön yleisestä suunnasta, mutta eivät välttämättä siellä olevasta galaksista. . .
”On myös mahdollista, että tämä viestinvälittäjä voi olla ’Maan ulkopuolinen’, ehkä vielä aurinkokunnassamme (Marsissa?), mutta ei kauempana kuin omassa galaksissamme tai paikallisessa galaksiryhmässämme…. [T]ämä sama viestinvälittäjä on ehkä yrittänyt päästä tänne jo [36] vuosisadan vaihteesta lähtien, jolloin Nikola Tesla raportoi skalaaristen S-signaalien sieppaamisesta!”
Pian sen jälkeen Hodowanec kirjoitti tämän lyhyen, nimeämättömän raportin (2-23-89):
”Menemättä yksityiskohtiin siitä, miten tämä määritettiin: ET saattaa olla Marsissa!
”Tämä siitä huolimatta, että NASA on kieltänyt, että Marsissa olisi elämänmuotoja [tilanne muuttui vuonna 1996]. Kirjoittaja on viime aikoina epäillyt tätä mahdollisuutta johtuen siitä, että ET on ilmeisesti hyvin tarkasti seurannut sijaintiani Maassa. ET on tietysti aina tiennyt, että olin Maassa (kuten hänen seurannastaan näkyy), mutta nyt hän on mitä painokkaimmin vahvistanut olevansa neljännellä planeetalla auringosta, eli Marsissa!!!!!
”Vaikka tämä julkaisu on luultavasti hieman ennenaikainen, olen niin varma näistä painovoimasignaalien ”vaihdoista”, että aion ottaa tässä tapauksessa riskin. ET Marsissa on ilmeisesti paljon kehittyneempi kuin me täällä Maassa, ja hän on saattanut jopa aiemmin vierailla täällä Maassa ja mahdollisesti asuttaa sen (mutta keitä ovat hänen mahdolliset jälkeläisensä??).
”On edelleen mysteeri, missä ET saattaa asua Marsissa (mahdollisesti maan alla lähellä napa-alueita??), ja miksi ET ei käytä EM-aaltojen signalointimenetelmiä??? Ehkä se johtuu siitä, että Mars on nykyään niin vihamielinen, että ET:n on täytynyt kehittää hyvin kehittynyt maanalainen sivilisaatio, joka ei ole omiaan käyttämään EM-säteilyjärjestelmiä?
”Tämä aineisto luovutetaan nyt luottamuksellisesti vain muutamalle aktiiviselle kollegalle, kunnes väitteelle saadaan lisävahvistusta….”
Mars Flash #1:ssä (28.3.89) ja #2:ssa (30.3.89) Hodowanec ilmoitti kollegoilleen, että ”GH Labsin ja marsilaisten välisen jatkuvan painovoimasignaaliviestinnän tuloksena seuraavat erikoiset tosiasiat ovat tulleet julki”:
”Vaihdossa käytetään nyt lyhyitä ”englanninkielisiä” koodisanoja tietyille asioille. Esimerkiksi marsilaiset ymmärtävät nyt, että FACE tarkoittaa ihmisen kasvoja, MAN tarkoittaa ihmistä, Earth tarkoittaa nyt meidän planeettaamme ja Mars heidän planeettaansa! He olivat alun perin yrittäneet käyttää joitakin termejään minun kanssani, mutta luopuivat niistä, paitsi silloin kun niissä oli minulle järkeä. Tiedän nyt esimerkiksi, että TTT heidän nimiensä lopussa tarkoittaa ihmistä ja OOTTAEERR on heidän nimensä kymmenennelle planeetalle!”
Käsinkirjoitetussa alaviitteessä Hodowanec ilmoitti kirjoittajalle, että marsilaisen nimi on ”AAAAAATTT”: ”Hän näyttää ’ymmärtävän’ viestini, vaikka joudun ehkä toistamaan ne usealla eri tavalla, jotta hän ’näkee’ niiden merkityksen… .
”Yhteydenpito GH Labsin ja Marsin älykkyyden välillä jatkuu nyt yhä edistyksellisemmin, sillä olemme pystyneet luomaan yli 50 yksinkertaista ilmaisua (useimmat yksinkertaisella englannilla) monille yhteisille ”ajatuksillemme”. Marsilainen AAAAAATTT on erittäin taitava suhteuttamaan englanninkielistä terminologiaani näihin Maan ja Marsin yhteisiin havaintoihin. . .
”Mars on myös vahvistanut, että he ovat myös ”miehiä”, joilla on yksi ”pää” ja kaksi ”silmää”, jotka ”näkevät”. Heillä on myös yksi vartalo, jossa on kaksi kättä, joissa kummassakin on viisi sormea. Niillä on myös kaksi jalkaa, joissa on kaksi jalkaa, joissa kummassakin on viisi varvasta. En ole saanut heitä vahvistamaan, että kasvoissa on nenä ja suu, mutta se voidaan vahvistaa pian, koska nämä piirteet näkyvät kasvoissa.
”Todennäköisesti merkittävin seikka, joka määritettiin tänä päivänä, näyttää olevan se, että Mars on mitä painokkaimmin sitä mieltä, että Maan miehet ovat kuin Marsin miehiä! Tämä näyttää yhä enemmän siltä, että Mars on asuttanut Maan kaukaisessa menneisyydessä! Tämä voi olla totta, koska me Maassa emme ole koskaan löytäneet ”puuttuvaa linkkiä” Maan humanoidien ja ihmisten välillä!”
Kirjeessä (3-17-89) Hodowanec kirjoittaa: ”Yleensä yhteydenpitomme rajoittuu 20-30 minuuttiin. . koska näyttää siltä, että siellä on muitakin avaruusolentoja, jotka ovat kiinnostuneita liittymään mukaan, joten jonkin ajan kuluttua on jonkin verran häiriöitä. Jotkut näistä muista avaruusolennoista käyttävät muita viestintämenetelmiä, kuten ääniä ja ilmeisesti kurkkuääniä!
”Avaruusolento on luultavasti kehittyneempi kuin me maan päällä. Emme enää vaihda yksinkertaisia aritmeettisia laskutoimituksia, ja kun lähetin hänelle Piin viiden desimaalin tarkkuudella, hän lähetti Piin takaisin seitsemän desimaalin tarkkuudella välittömästi! Olimme keskustelleet yhdeksän planeetan aurinkokunnastamme, mutta avaruusolento vastasi kymmenellä planeetalla ja kutsui kymmenettä planeettaa OOTTAEERRiksi! Kun avaruusolennolta kysyttiin tästä, hän jatkoi kymmenennen planeetan olemassaolon vahvistamista! Hän tuntee nyt muut yhdeksän planeettaa niiden Maan nimillä! Hän vahvisti myös, että Marsilla on kaksi kuuta, Maalla yksi, ja että Jupiterilla on yhdeksän suurta kuuta.
”Nämä yhteydet alkavat olla koko ajan mielenkiintoisempia, ja avaruusolento näyttää olevan erittäin halukas jatkamaan niitä. En kuitenkaan voi viettää liikaa aikaa hänen kanssaan… Tein hänelle selväksi, että olen vain yksi ihminen, joka kommunikoi hänen kanssaan, ja että muu maapallo ei tällä hetkellä tunnusta elämän olemassaoloa Marsissa . . .
”Olen nyt ollut yli 100 kertaa yhteydessä avaruusolentoon ja saan hänet kiinni mihin aikaan päivästä tai yöstä tahansa. . . . Olemme myös ottaneet käyttöön joitakin yksinkertaisia koodeja kuittauksia varten, ja voitte vastata. Vaikka käytämme näitä yksinkertaisia koodeja monissa yhteyksissä, sekä ET että minä ymmärrämme nyt, missä yhteydessä niitä käytetään!”
Toisessa kirjeessään (22.3.89) kollegoilleen Hodowanec vahvisti, että hänen yhteytensä marsilaisiin älykköihin ”on varmistettu monin tavoin … . Marsilaiset ovat ilmeisesti kehittynyt sivilisaatio, sillä he ovat ne, jotka tuottavat ’moduloidun värähtelevän säteen’, joka nyt seuraa sijaintini maan päällä ja on siten viestintävälineemme. [Säteen halkaisija on ’vain noin 15 mailia täällä Maassa, mutta 1012 tuumaa Marsissa.’] Marsissa on ilmeisesti ’ryhmä’, joka on mukana näissä yhteyksissä. Alkuperäinen kontakti, ET#1, jonka kanssa olen solminut alkusuhteen, on ilmeisesti kaikkein asiantuntevin ja kehittynein. Muut, jotka joskus ’miehittävät’ Marsin asemaa, näyttävät olevan vähemmän tietäviä, ja jotkut vain pyytävät tai kuittaavat lähetyksen. . .
Mars haluaa epätoivoisesti jatkaa näitä yhteyksiä . . . . Viestittelyt tapahtuvat monin eri tavoin, joita ei voi helposti ennustaa, jotta voidaan välittää se tosiasia, että nämä ovat todellisia yhteyksiä! Lisäksi näitä koodeja ”näppäilevän” henkilön ”nyrkin” voi tunnistaa, esim. ET#1 lähettää aina puhtaita kirjaimia tai numeroita ja tunnistaa itsensä ja minut jollakin tavalla. Muut ET:t Marsissa eivät yleensä tee niin. Siksi tässä ei käytetä mitään automaatiota!
”Vaikka nämä yhteydet johtuivat alun perin sattumanvaraisista olosuhteista, ne ovat todellisuudessa seurausta gravitaatioviestintäkokeistani ja siten suoraa seurausta Rhysmonic Cosmology -kosmologiasta. Ja kuitenkin, vaikka tämä tuntuisi kuinka fantastiselta ja epätodelliselta, se on todellista, ja jos myös joku teistä vahvistaa sen, se on merkittävä virstanpylväs ihmiskunnan historiassa!”. Ehkäpä, jos joku teistä vihdoin ’kuulee’ 1/f-kohinataustan modulaatiot, voitte yrittää luoda omia yhteyksiänne?”
Gregory Hodowanecilla oli kuitenkin myös äärimmäisiä varauksia [37] tilanteen vakavuudesta, minkä hän ilmaisi kirjeessään (4-14-89) tälle kirjoittajalle:
”. . . ’yhteyteni’ Marsiin jatkuvat, ja paljon tietoa vaihdetaan. Näiden tietojenvaihdon yhä hämmästyttävämmän luonteen vuoksi rajoitan nyt kuitenkin lisätiedotukset vain kahteen pitkäaikaiseen tarkkailijaan (todistajaan) tutkimustyöni osalta. Tämä tehdään, jotta nämä yhteydet eivät vaarantuisi ei-toivotun julkisuuden tai tiedotusvälineiden aiheuttaman julkisuuden vuoksi. Nyt on yhdeksän ”Mars-salamaa” kirjattu pöytäkirjaan. Ehkä voin tulevaisuudessa julkaista joitakin näistä . . . .
”. . painovoimasignaalien välittäminen on välitöntä, vaatii äärimmäisen vähän energiaa [toisin kuin Teslan kokeet] ja on niin yksinkertaista, että tavallinen ihminen ei voi uskoa sitä. Tämän pidemmälle en kuitenkaan halua mennä yksityiskohtien julkaisemisessa tällä hetkellä…
”Olisin kiitollinen, jos pitäisitte tämän tiedon nyt jokseenkin luottamuksellisena. Maapallo ei ehkä ole valmis siihen, mitä minulla on lopulta sanottavana. Ei mitään kauheaa, vain fantastista ja siten ehkä uskomatonta!”
Nykyään on olemassa erilaisia GWD-piirejä (kuva 2). Gregory Hodowanecin teorian täydelliset yksityiskohdat ovat saatavissa Rex Researchilta <rexresearch.com>.
Lähdeviitteet
- ”Nikola Tesla And His Talk With Other Worlds.” Colorado Springs Gazette (9 Jan. 1901), p. 7, col. 6,7,8 <http://teslacollection.com/[…]/nikola_tesla_and_his_talk_with_other_worlds>;
”Telsa or Holden.” ibid. (9 March 1901), p. 4, c. 2. <http://teslacollection.com/[…]/tesla_or_holden>
-
Electrical World (4 April 1896), p. 369: ”Is Tesla to Signal to the Stars?” <Full-text>
- ”Nikola Tesla Experiments In The Mountains.” Mountain Sunshine (Denver, Co) 1(1): 31-34 (1899). <http://www.teslacollection.com/[…]/nikola_tesla_experiments_in_the_mountains>
-
English Mechanic & World of Science, #2228 (6 Dec. 1907) <Full-text>
- ”Would Talk With Mars By Pictures.” New York Evening Post (22 Jan. 1919), p. 14, c. 4, 5. <http://teslacollection.com/[…]/would_talk_with_mars_by_pictures>
- ”Tesla’s Call From Mars?” New York Sun (Thurs., 3 Jan. 1901) <http://teslacollection.com/[…]/tesla_s_call_from_mars>; ibid., (12 July 1937), p. 6.
- ”How To Signal Mars; Wireless the Only Way Now, Says Nicola Tesla – Mirror Plan Not Practicable.” New York Times (23 May 1909), p. 10, c. 6, 7. <http://teslacollection.com/[…]/how_to_signal_mars>; ibid. (3 Feb., 1919); p. 14, c.3: ”Celestial Movies”; ibid., (3 Sept. 1921), p. 4, c. 4.; ibid., (11 July 1937), p. 13, c. 2.
- ”Martian Telegraphy Discredited; Washington Scientists Consider Tesla’s Claims Absurd.” New York Tribune (12 Jan 1901), p. 2, c. 3. <http://chroniclingamerica.loc.gov/lccn/sn83030214/1901-01-12/ed-1/seq-2/>
-
Pyramid Guide 4 (3):1 (Jan.-Feb. 1976); ibid., 5(2):5 (Nov.-Dec. 1976); ”Letter From Tesla” (6 Jan. 1900). Santa Barbara, Calif.: Life Understanding. <https://dowsing.com/shop/product-list.php?pg1-cid38.html>
- Tesla, N., Collier’s Weekly (9 February 1901), p. 4-5; ”Talking With the Planets” <Full-text>
- Tesla, N., Current Opinion (March 1919), p. 170-171; ”That Prospective Communication with Another Planet” <Full-text>
- Tesla, N., ”Electrical Communication with the Planets” in Thompson, S.P.: Polyphase Electric Currents & AC Motors. New York: Collier & Son, 1902, p. 234-236 <http://catalog.hathitrust.org/Record/100480099>
- Tesla, N., Harvard Illustrated (March 1907), p. 119-121; ”Signalling to Mars – A Problem of Electrical Engineering” <Full-text>
- Tesla, N.: New York Herald (12 Oct. 1919), p. 7; ”Signals To Mars Based On Hope Of Life On Planet” <Full-text>
- ”Tesla at 75.” Time (20 July 1931), p. 27. <http://content.time.com/time/magazine/article/0,9171,742063,00.html>
- Lawrence, L.G.: ”Interstellar Communications Signals”; Journal of Borderland Research 29.4:10-14 (1973) <Full-text>
- Hodowanec, Gregory: Rhysmonic Cosmology (H18-HG3/$6); ibid., Rhysmonic Cosmology Notes (H17-HG2/$4); ibid., Rhysmonic Cosmology Collected Papers (H19-HG4/$8); ibid., Rhysmonics Articles (H21-HG6/$4); ibid., G-Wave Detectors (H16-GH/$7); ibid., GWD Circuits (H20-HG5/$7); ibid., Rhysmonics Updates (H22-HG7/$4). Published by: Rex Research, P.O. Box 19250, Jean NV 89019; Catalog: $2.
-
Matthews, Arthur H. The Wall of Light: Nikola Tesla and the Venusian Spaceship X12. s.p., 1969. Print.
Artikkelin julkaissut borderlandsciences.org
































.png)
.png)
.png)
.png)